Life Advice, Neuroscience, Personal Perspective

As we approach the end of the calendar year and are in the midst of the holiday season, time is often top of mind. Both reminiscing on the year that was or holidays past are common in December. Memories often made with those we love can bring comfort if we are apart or if those individuals are no longer with us. We can mentally time travel back to the past and in the process drag the positive feelings from that time into the present, if only for a bit.
In addition, as the calendar transitions to January many people begin to think about the new year and our goals for the future. We project ourselves into the future and ask: what can we do now and in the immediate months ahead to get us where we aspire to be years from now? Getting more "in shape" today will make us healthier long-term, saving a bit more for retirement now will pay off when we are older, learning a new skill over the next few months can make us more marketable for a future career. These are all worthy goals that involve simulating how our actions in the present will potentially impact our future.
In both instances mentioned above we are constructing other moments in time in our heads and using them to affect our current state or guide near-term actions. Thinking about both the past and future also reminds us that the passage of time is constant and persistent and that the present is quite fleeting. The clock ticks on around us pushing the present into "the past" and pulling "the future" into the present. And while we are certainly aware that time is a real, objective thing that can be measured in second, hours, days, and years it is also, in a way, a subjective experience that can be hard to fully describe.

Measuring Time as a Mortal
Time is certainly an intriguing concept, especially to human beings. We have all experienced time seeming to slow down or speed up in our daily lives. In some ways it can seem arbitrary (who decided a minute is 60 seconds?) but in other ways it is very tangible and real, relentlessly moving forward (regardless of how you measure it) and in the process reminding us that it is personally limited. Time is our most precious resource as it is finite. You cannot create more of it. Objectively, time is constant and immutable in the sense that one cannot escape it.
"To be human is to be aware of the passage of time; no concept lies closer to the core of our consciousness."
- Dan Falk from In Search of Time: The History, Physics, and Philosophy of Time
- Dan Falk from In Search of Time: The History, Physics, and Philosophy of Time
Our fascination with and fixation on time is most certainly related to our mortality in that we all will die in time. All living things have an expiration date and one could reasonably argue that the act of living is the delaying of death through information and action.
Time as our most precious commodity can perhaps best illustrated by the fact that some of the world's most wealthy and powerful people have been working hard to extend the time they have on Earth with investments in Altos Labs, Unity Biotechnology, Juvenescence, and Calico Labs and organizations like the Methuselah Foundation. It has become popular these days for many of us to try to hack our longevity (for more see the work of Peter Attia including his podcast and new book Outlive: The Science & Art of Longevity for more on this). And while certainly having information and resources could increase one's longevity and potentially extend one's time on Earth, time as a daily commodity is the same for everyone.
We all have 24 hours in our days which we fill with commitments (work, raising a family), biological functions (eating, sleeping), and exercising our own preferences and desires. And while certainly those with means and privilege may be able to spend less hours on "basic needs", they can't extract more than 24 hours out of a day. Those 24 hours are limited and precious for everyone. So, what will you do with the 24 hours you have each day? Should you seek to optimize them to the hilt (to try to squeeze all utility out of them) or is their value in "wasting time" or enjoying your time? And how should you balance the allocation of these activities?

Structuring Time
The quote "How we spend our days is how we spend our lives" is attributed to the author Annie Dillard in her book The Writing Life. She goes on to speak to the value of daily schedules defending us from "chaos and whim" and that "a schedule is a mock-up of reason and order—willed, faked, and so brought into being; it is a peace and a haven set into the wreck of time; it is a lifeboat on which you find yourself, decades later, still living."
This quote speaks to both the unpredictability of the future and inability for us to completely "master time" (chaos and whim; the wreck of time) while also conveying the value of having structure in our lives to ensure we make time for what is important to us. This could include key work priorities or making time for companionship and fun, all of which are important to human flourishing. But in our limited time, how do we prioritize amongst the many things we could be doing while also ensuring we make progress on what we have to do to survive? There are no easy answers. Rather, we must try to the best of our human ability to plan for the future while not letting the present pass us by.
The Burden of the Present: A never-ending to-do list
Given that most of us perceive we lack sufficient time in our daily lives, many search (often unsuccessfully) for various approaches to boost their productivity and "maximize their time." The rise of smart phones and communication technology like Slack and Zoom make it possible for us to work from virtually anywhere at anytime. It is exceedingly difficult to disconnect in such an environment. I think many of us believed, especially with the rise of "remote work" in response to the global COVID-19 pandemic in Spring 2020 that we would be liberated from the bounds of an office, strict working schedules, and the dreadful daily commute. And in a way we were but when the lines between work and home blur it makes separating one's "work time" from "personal time" a challenge. Even the most exciting sounding possibility for remote work technology - work from anywhere, including while on vacation - seems to miss the point that one takes a vacation to, at least conceptually, escape work.
There are plenty of resources and writings that try to address time management in the digital age or successfully setting and maintaining work/life boundaries. Various tools and apps promise to help. And while certainly there may be some value in these approaches, I think we have all found a "solution" to our time management problems elusive. So did author Oliver Burkeman who, in his book Four Thousand Week: Time Management for Mortals, comes to a shocking but perhaps accurate conclusion that we have more responsibilities and work to be done than can ever be accomplished in our lifetime (the average length of which is 4,000 weeks). He says we should accept this reality and in the process try to liberate ourselves from the notion that there is some hack or approach that will help us accomplish our many tasks. When I read this book, its central thesis shocked me at first. I expected to get some practical tips or insights on time management and was left with the hard truth of the inability to ever "accomplish my work". This is something I am still coming to terms with even though at my core I realize it is, in fact, true.
| In the book, Burkeman makes the point that we often want to rationalize that the hard work we do today will pay off in the future. And there is some truth to this. But, he also mentions we wrongly believe that through our present efforts and sacrifices - the checking off of our "to-do list" - we will ultimately reach some idealized future where we will magically "have more time" and then be able to focus on what really matters. |  |
However, there are always new things we will (or feel we) need to do...the to-do list is essentially never able to be fully checked off. The challenge we all encounter is the idealized future we are "working toward" is often a future that we never pull into the present...it never reaches us.
So, why be so worried about all these responsibilities and to-dos if they prevent us from living in the now? Why not just live fully in the present - YOLO and all of that? Well, living too much in the present, the NOW, also has its issues.
The Neuroscience of Time: NOW vs LATER Rewards, Depression, & Anxiety
A quick detour into how our brains perceive time. First, there is very interesting work from Thomas Suddendorf and Michael Corballis suggesting that "mental time travel" (ie, the ability to transport our consciousness to the past - episodic memory - or future - prospection) may make humans unique or at least is a process more developed in humans. These researchers also speculate that human language may have evolved to facilitate group mental time travel through stories from the past and/or coordinating/planning future, collective goals.
So, our brains are built for time traveling. They also can get fixated at different points in time to our peril - ruminating too much on the past and/or becoming too anxious about the "unpredictable" future. We also struggle calculating the time value of money and are rightly biased to be more concerned about having a resource like money to spend today versus investing it for a future purpose.
Living wholly in the present has its appeal and deep biological origins in our animalistic need for survival. Humans and other animals indeed will "discount the future," favoring rewards available NOW (or soon) over those delivered in the future, even if the future rewards are large. Academics call this tendency to favor NOW over LATER delay discounting behavior.
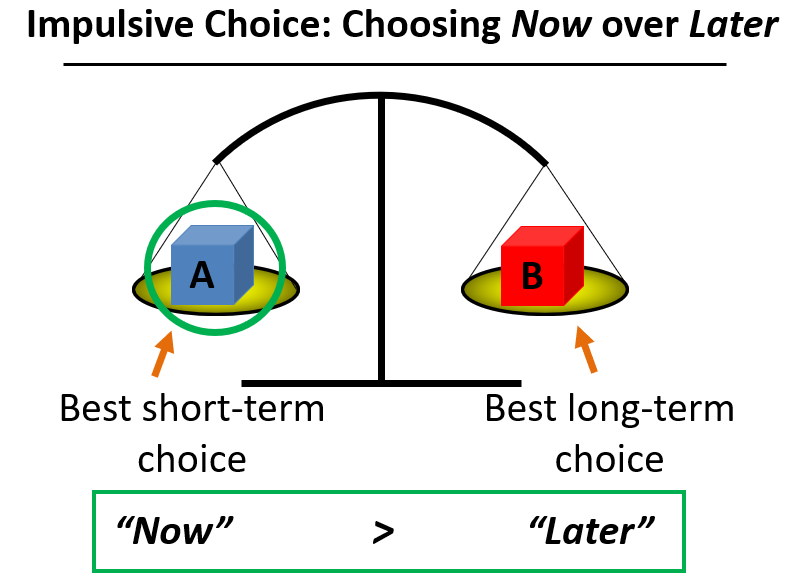 | 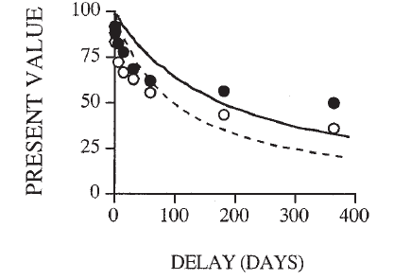 |
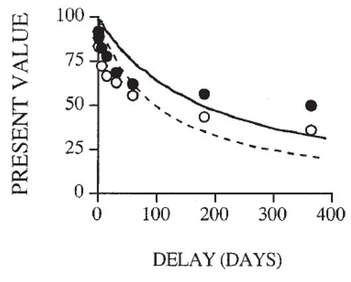
The present value of a reward is discounted in proportion to the time you must wait to claim it. The figure on the left shows how impulsive choice is a preference to choose the NOW option over the LATER option at higher rates than average or, to put it another way, to discount the future more steeply. In the figure on the right we see two "discounting curves" where the white circles reflect an individual with a steeper discounting rate (ie, they value the NOW over the LATER more or discount the future more steeply) than the individual plotted with black circles.
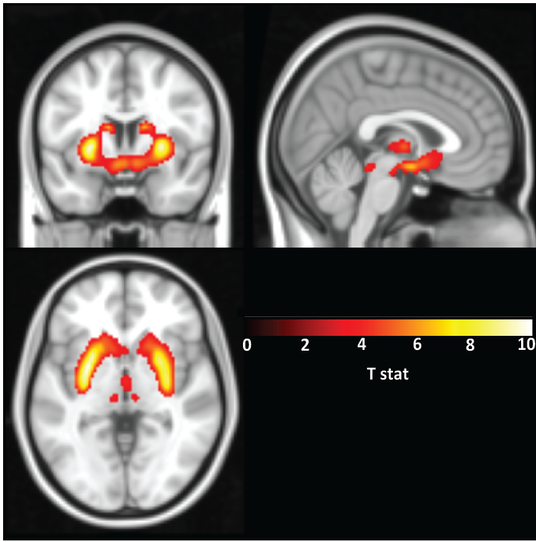
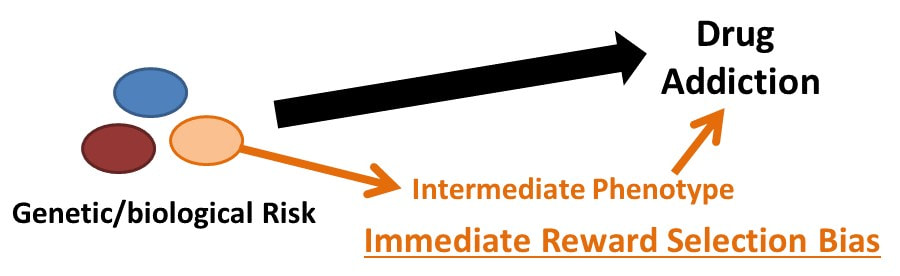
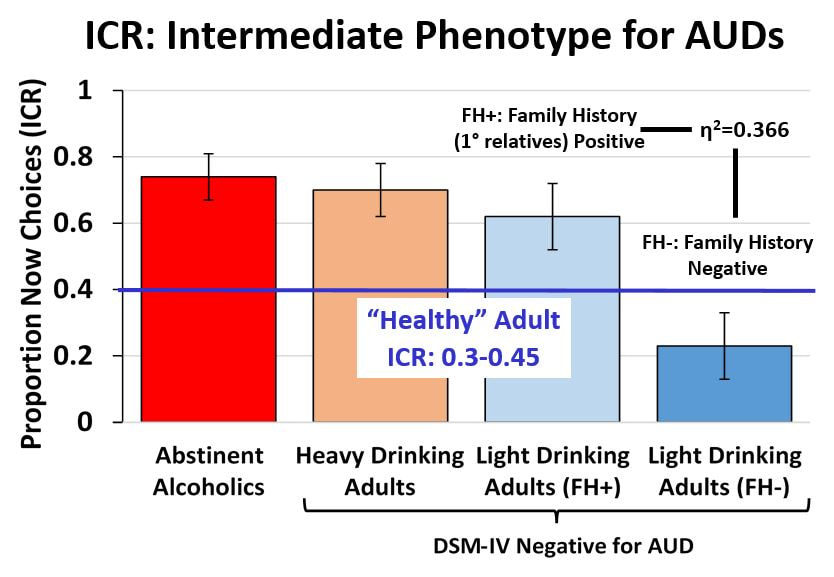
In short, we are biologically biased to favor the NOW over LATER, though some of us more than others. My Ph.D. dissertation work at the University of North Carolina was focused on investigating these individual differences in delay discounting behavior in humans. As mentioned, we know from years or research on a variety of species that rewards have less value if they are delayed and the degree to which individuals discount this delay varies. In humans, these variations in delay discounting are based partially on biology and our environment but can also be susceptible to temporary states such as acute stress. And understanding this phenomenon is important as delay discounting has also been shown to be associated with drug addiction and may indeed be a risk factor in developing addictive behaviors.
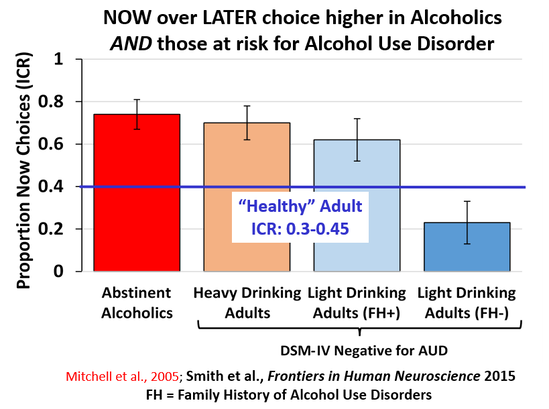
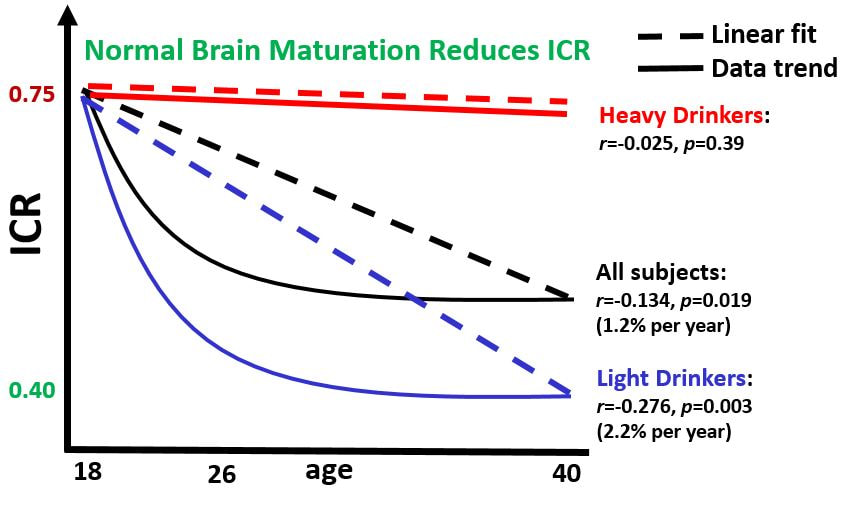
My dissertation work found that individuals at risk for alcohol use disorders (heavy drinkers, those with a family history of alcohol use disorders) show a higher preference for NOW choices than those at low risk (no family history, light drinking behavior).
While certainly we can see how thinking only about the NOW and our impulsive urges may be problematic, so too is an over-emphasis of our thoughts on the past or future. It has been demonstrated that depression is more associated with past events while anxiety more often associates with future events. This indicates that an unusual fixation on both the past and future can also be problematic to our mental health and well-being.
Thus, balancing our focus on past, present, and future is probably best to live an effective life. One could see how learning from the past can enhance future survival and similarly how planning for the future (and the uncertainty of future events) is also important. And living in the present is also essential as it is the only actual living we are doing currently.
The cliché phrase (attributed to Eleanor Roosevelt, among others):
Yesterday is history. Tomorrow is a mystery. Today is a gift, that is why they call it the Present.
May by a bit trite but that doesn't mean there is not wisdom in it.
It speaks to the idea that the present is within our control as it is happening now.
Yesterday is history. Tomorrow is a mystery. Today is a gift, that is why they call it the Present.
May by a bit trite but that doesn't mean there is not wisdom in it.
It speaks to the idea that the present is within our control as it is happening now.
Focusing too much on the past or future prevents us from living in the moment and taking actual action in the here and now.
Living in the Present
In some ways we think living in the present moment (right now) should be easy. However, to be really present in a moment requires you to resist distraction. Sitting with your thoughts or being engaged in the presence of others (ie, having meaningful conversations and connections) can be surprisingly uncomfortable if you haven't done it frequently.
In an age of unlimited work and entertainment distractions, one of the most difficult things for us is to be truly present in a moment. Part of the challenge is that we know at a deep level that our time is finite and limited and so we often attempt to master it, to maximize it. Sitting in the moment and enjoying it can seem somehow wasteful and superfluous - what do we get from being "in the moment"? Well, being present in the moment is effectively at the center of the practice of mindfulness, which has shown a range of positive effects on human health and well-being. Despite this knowledge that being in the moment is valuable, we almost can't help our thoughts from drifting to our "to-do list" or the need to use our time "better".
At the center of our focus on optimizing time is the realization that we are mortal and finite...that we will die. Time will run out for all of us. The irony of this fact, though, is that when we accept this, really accept this we can stop trying to convince ourselves that by maximizing our schedules and time we will somehow create more of it. Rather, realizing our mortality and relishing the present can help us focus on what matter most to us and stop waiting for "one day" to come.
An excellent example of the "waiting for one day" phenomenon is the fixation many Americans have on earning and saving more money today, often at great personal sacrifice, for that glorious retirement in the future.

Preparing for the Future
In the personal finance realm, a focus on saving for the future is seen as an unequivocally good thing. Indeed saving for the future is critical to retiring in a financially stable position (especially if your government does not provide a strong social safety net to retirees). However, it is very easy for many individuals, especially those obsessed with the Financial Independence, Retire Early (FIRE) movement to overly focus on future retirement instead of present satisfaction. Many individuals have gone to great lengths to cut so much out of their current lives to expand their savings rate that they live lives of immense deprivation (NOTE: there is a difference between frugality and deprivation).
But one should reflect during their working lives and ask: What are they saving this money for? What is the purpose of having retirement savings? Upon deeper self-reflection you could come to the conclusion that there is such a thing as delaying one's consumption too much into the future.
Also, as none of us really know how long we will live, we could be saving for a future that may not come. The influential book Die With Zero may have an extreme title but its thesis seems correct to me: one should think critically about when it makes sense to spend money NOW on experiences that should not be delayed for a future retirement.
In the book, the author speaks to the need for us to consider when we should have certain experiences and questions whether waiting "until retirement" is the right answer. Backpacking through Europe is probably best done in your twenties, for instance. Even something like hiking a beautiful trail may be more feasible when you are forty or fifty not seventy. And the thought experiment goes even further when it brings up the point that you can only do certain activities with others during select windows of time. You can't constantly put off playing baseball with your 10-year old because they won't be 10 for more than 365 days. You can't keep delaying that big family trip with your parents until "your work slows down" because their time on Earth is probably more limited than yours (and the work will probably never "slow down"). When you bring other people and their mortality into the equation or, less grimly, the fact that they are also aging and moving into different seasons of life themselves, you reach the conclusion that you can't afford to wait to do certain things with them. This doesn't have to be a morbid or depressing experience (ie, focusing on a loved one's aging and mortality) but hopefully it is a motivating force to take action NOW to have memorable experiences with the people important to you.
Memory Dividends
These memorable experiences produce what the author of Die With Zero calls "memory dividends". Essentially, these experiences with others live on in our memories for years and the recalling of them can bring joy and fulfillment well into the future. So, by investing the time in key experiences and moments NOW we reap the benefits in the future via our memories and recollection of "that time when" story. This is not unlike investing for retirement and reaping dividends on our capital...we put money in now for a payoff later. But in the case of memory dividends we often sacrifice money now for experiences we will treasure forever. Money can't buy happiness but it can provide us with the means to have special experiences with others we will treasure.
| And while a lot of this memory dividend talk brings to mind spending lavishly on a multi-thousand dollar African Safari, Mediterranean Cruise, or week-long trip to Disney World, one doesn't have to spend a lot of money to make memories with family and friends. In a way, being present with those we care about can be enough. Making the time for them and prioritizing that over other commitments can be everything. Ironically, we often remember some of the smallest, seemingly mundane experiences with those we love as strongly as the big, audacious trips and outings. And these memories often become more powerful with the passing of time and, ultimately, sadly, when those we made them with are no longer with us. Then, the memory is what is left in the end. A reminder of that individual and their impact on you and you on them. |  |
Nostalgia is defined as a sentimental longing or wistful affection for the past. For example, we often look back on our times as children fondly when we perceive life was simpler and less stressful. And data show older adults are more likely to be nostalgic as they riminess on their youth or days gone by and in general these nostalgic experiences lead to higher subjective well-being in the elderly. So, in a way, nostalgia and positively remembering the past allows us to cope with approaching the inevitability of death as we age. It seems a bit morbid at first but our brains and consciousness are probably doing us a big favor by increasing our subjective well-being as we age. It often helps us feel we had a good life.
Immortality
At the beginning of this piece I mentioned the fixation many of the world's billionaires have on living longer (or perhaps forever). And while advances in technology may one day allow those with means to upload their consciousness to the cloud or a robot body (saying nothing about the ethics of whether they should do it), what if I told you there is something you can do today to ensure your immortality?
We have already covered this secret. Your existence, your impact, your story can live on in others. When we interact with those around us we are laying the groundwork for a sort of oral history of ourselves. How we treat them will often be recounted. The person we were in their eyes can persist for years to come. So, what kind of person do you want to be remembered as?

This brings to mind the concept of "resume virtues" and "eulogy virtues" from David Brooks. Brooks argues that what we all should be focused on are the actions that could end up in a funeral eulogy (he was a kind, caring husband and father who gave much of his time to his favorite charity after retirement) and not points on a resume (he rose to the rank of senior vice president). In the moment, however, we often are more fixated on the resume items. It is up to us to reflect often on our actions and ask ourselves if the things we are obsessed by in the now will be things we and others care about in the future.
We build our eulogy virtues through caring for others.
And, lest we forget, if you are a creative, writer, or scientist, we also build our legacy through our work. So, doing good work is important in the sense that if it produces a piece of knowledge or creative work that others can consume, be inspired by, and build on and through this process has the potential to live on after us. And sometimes what we put out there in the world can have an effect or impact we can't even imagine (or experience) during our life. For example, in the sciences there are many who question the value of "basic research" but this type of work can inform big breakthroughs, often years later (see for example, Mendelian genetics, the decades-long development of technology for mRNA vaccines, and an overview of other innovations from fundamental research published by the National Academies of Science, Engineering, and Medicine). I say all this to emphasize that anything you put out in the world, from a product of your creative or cognitive labor to the way you treat others, can have long-lasting and unexpected effects. The key is to do these things without always questioning (or obsessing over) their "value" in the now. I know that is easier said than done but history shows underappreciated work can have a big impact later.

And while "history" may not chronicle the impact good deeds and kindness have on others at an individual level we all know at our core that how others treat us affects us. Think back to an elementary school teacher in your life...they almost certainly positively impacted you in ways they may never know. And they do the work because they care and aren't necessarily focused on "measuring" or "tracking" the product of their labor. In the immediate term there are crude and ineffective ways to "measure" a teacher's impact (ie, test scores) but in the long term their impact is immeasurable (igniting curiosity in young people, encouraging a student to pursue a passion, or modeling how to be a caring citizen). Measuring one's impact on others is hard unless they tell you explicitly about it but that doesn't mean an impact wasn't made.
In the end, all of you reading this (at least with the current state of technology in late 2023) will eventually move on to whatever comes after life. As such, your life is precious and limited. So, how will you spend it?
In my mind, our jobs as living beings is to try our best to bring joy, happiness, and meaning to ourselves and others while also adding value to the world. Sometimes the value you add is obvious, sometimes less so. In a way, it is not for you to judge or measure your value (though I know we all do it...hey, we are human beings after all). But if you deeply examine how you can take actions or deploy your skills to serve others today and in the future, whether they be family, friends, or your fellow man, I promise you that you will have made good use of your limited time on Earth.
More from the blog:
For Further Reading:
Watch:
- The power of human connection
- Why you should get involved in things outside the lab/work
- Cultivate serendipity by giving back and getting involved
- Find your passion? Finding meaning and purpose in your work and life
For Further Reading:
- The Science of Mental Time Travel and Why Our Ability to Imagine the Future Is Essential to Our Humanity
- In Search of Time: The History, Physics, and Philosophy of Time (Book)
- Your Brain is a Time Machine (Book)
- Die With Zero (Book)
Watch:







 RSS Feed
RSS Feed